30 de diciembre de 2020
Entrenamiento de fuerza y ciencia
Potenciación post-activación: mecanismos, aplicaciones y limitaciones.
El ejercicio puede considerarse como un tipo de estresor que, como cualquier otro, es capaz de ejercer una respuesta biológica (o respuesta al estrés). Una característica común de todos los estresores es que hay una fase aguda, durante la que se producen ajustes homeostáticos; una fase más crónica, durante la cual el estresor puede ser acomodado por las adaptaciones; y, por último, una fase de agotamiento, durante la cual se producen inadaptaciones.
Los científicos que estudian el deporte, han estado obsesionados históricamente con la respuesta crónica a la perturbación homeostática mediada por el ejercicio (también conocido como adaptación) como la hipertrofia. Sin embargo, la contracción muscular es capaz de provocar una gran variedad de respuestas homeostáticas agudas, como la alteración del flujo sanguíneo a los músculos activos; el aumento del ritmo cardíaco; el aumento del ritmo respiratorio; el aumento del consumo de oxígeno; el aumento de sudoración; el aumento de la temperatura corporal; la secreción de hormonas del estrés como la hormona adrenocorticotrópica (ACTH), el cortisol y las catecolaminas; el aumento del flujo glicolítico; y la alteración del reclutamiento de los músculos. Estos cambios son transitorios y vuelven a los niveles de referencia después del ejercicio1.
Nosotros, como profesionales de la fuerza y el acondicionamiento, estamos constantemente buscando e intentando implementar una variedad de métodos de entrenamiento para mejorar el rendimiento de nuestro atleta, no sólo de forma crónica, sino también aguda. Cada vez que nos cuesta encontrar un protocolo de calentamiento puntero para mejorar una sesión de entrenamiento o el rendimiento de un día de partido, en el fondo sólo estamos tratando de manipular cada uno de los procesos fisiológicos con características de mejora para nuestro beneficio.
Una técnica de entrenamiento de fuerza que se ha convertido en objeto de investigación reciente y de interés para entrenadores y atletas es la potenciación post-activación (PAP), un fenómeno por el cual la función muscular se incrementa como resultado directo de su historia contráctil2. La PAP es el proceso fisiológico en el que se basa el entrenamiento complejo y de contraste; ambas metodologías se desarrollaron para inducir un estado muscular de «preparación» o potenciación.
La PAP se ha definido como una respuesta muscular contráctil mejorada para un nivel determinado de estimulación tras una contracción voluntaria intensa, que se mide como la fuerza máxima de contracción provocada por la estimulación eléctrica supramáxima3. Sin embargo, en la literatura se han descrito muchas formas de potenciación que se suelen denominar erróneamente PAP. Lo más probable es que no se esté midiendo o deduciendo realmente la PAP (que implica la confirmación mediante estimulaciones de espasmos) sino las mejoras del rendimiento de la potenciación posterior a la activación (PAPE), que es básicamente la fuerza voluntaria o la mejora de la potencia que se observa después de un calentamiento de alta intensidad basado en el ejercicio y que se atribuye a otros factores que influyen en la función muscular (por ejemplo, la temperatura del músculo, el nivel de activación/aprendizaje) .
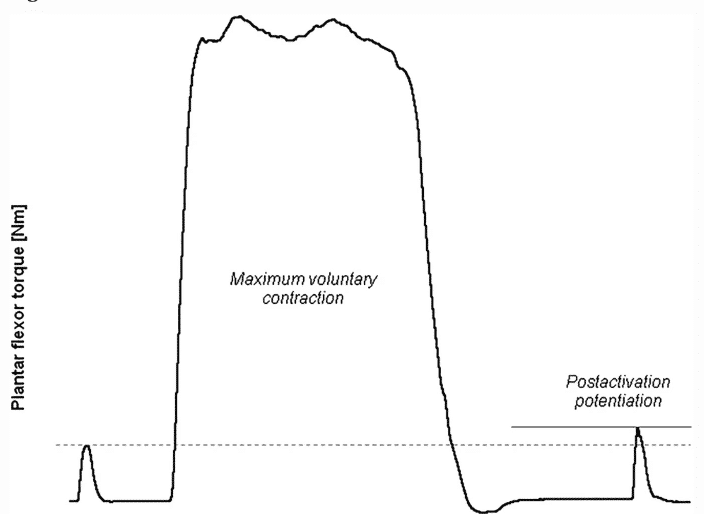
Figura 1 Ejemplo de la medición de la potenciación post-activación (PAP) en los músculos flexores plantares. Se provoca artificialmente una contracción de base en los flexores plantares en reposo. Dos segundos después de una contracción voluntaria máxima condicionada, el espasmo provocado tiene un par máximo mayor en comparación con el tic de la línea de base. El incremento del par máximo de contracción de la línea de base (línea discontinua) al par máximo de contracción posterior a la contracción (línea continua) corresponde a la extensión del PAP5
Breve Historia y evolución de la terminología
Los primeros trabajos sobre la capacidad potenciadora de ciertas teorías musculares datan de los inicios de la fisiología del ejercicio en el siglo XIX. El término más antiguo utilizado en la literatura parece ser «treppe» (escalera) en el artículo «La causa del treppe «6 de 1906, que se refiere a las respuestas repetidas de un tejido determinado al aumento repetido e igual de los estímulos durante un tiempo en intensidad. Aunque el concepto no era nuevo en ese momento, se originó en la innovadora labor realizada por investigadores alemanes en 1865 y 18717.
Desde entonces, se han descrito en la literatura tres formas diferentes de potenciación: escalera, potenciación postetánica y PAP4. Todas ellas tienen una característica común: la potenciación de la respuesta contráctil tras la aplicación de un estímulo condicionante. Además, la principal diferencia entre las tres formas de potenciación es el estímulo condicionante; en la escalera, las contracciones condicionantes comprenden estímulos eléctricos repetidos de baja frecuencia en los que las contracciones aumentan secuencialmente en amplitud; en la potenciación postetánica, un breve tren de estimulación eléctrica de alta frecuencia actúa como contracción condicionante8,9.; en el PAP, el aumento de la respuesta contráctil (contracción muscular) se evoca mediante la activación muscular voluntaria4,10.
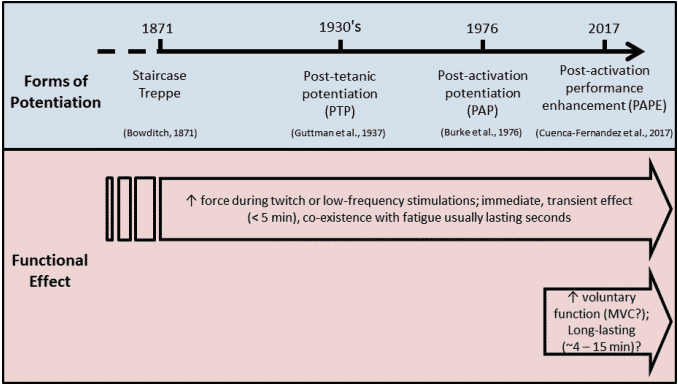
Figura 2 Muchas formas de potenciación han sido descritas en la literatura desde el siglo XIX4
Durante los últimos 50 años, los investigadores han estado estudiando el PAP en diferentes tipos de sujetos, condiciones y metodologías, haciendo grandes progresos hacia una mejor comprensión de sus mecanismos, implicaciones y mejores prácticas. Sin embargo, dado que para una evaluación adecuada de la PAP es obligatorio incorporar un espasmo provocado eléctricamente o una estimulación tetánica de baja frecuencia para determinar su magnitud en el punto de la prueba muscular voluntaria, así como la gestión adecuada del curso temporal del efecto de la PAP, no siempre está claro si la PAP «clásica» sustenta la mejora del rendimiento4. Además, se ha formulado la hipótesis de que la base de estas mejoras del rendimiento in vivo con un curso temporal diferente puede deberse a un mecanismo diferente. Así pues, se ha propuesto que se utilice la expresión «mejora del rendimiento posterior a la activación» (PAPE) cuando se realicen contracción(es) de acondicionamiento voluntario de alta intensidad con la intención de mejorar la producción de fuerza voluntaria posterior, en lugar de la provocada eléctricamente (twitch) por la producción de fuerza4.
Mecanismo
Antes de empezar a hablar del mecanismo fisiológico responsable del fenómeno fisiológico del PAP, es importante describir la contracción muscular a nivel molecular.
Para que se produzca la contracción muscular, el potencial eléctrico, generado en la neurona motora, tiene que penetrar en la maquinaria contráctil del músculo, esto se consigue transmitiendo los potenciales junto con el sistema de túbulos transversales (túbulos T). Los receptores de dihidropiridina sensibles al voltaje situados en los túbulos T se activan por la despolarización de la membrana inducida por el potencial de acción y, por lo tanto, interactúan directamente con el receptor de ryanodina (Ca2 + canal de liberación) situado en el retículo sarcoplásmico. Posteriormente, la despolarización de la membrana induce la liberación del Ca2 + contenido en el retículo sarcoplasmático en el espacio adyacente a las miofibrillas y activa los mecanismos contráctiles11,12.
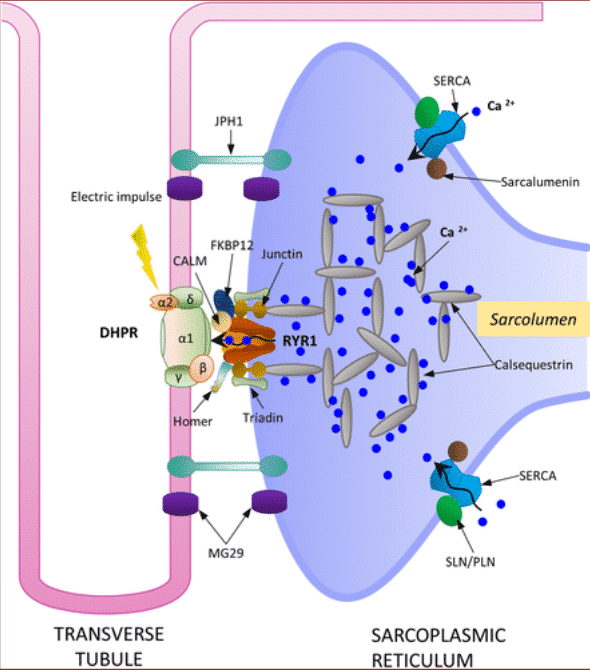
Figura 3 Estructura y función de los túbulos T11.
Cuando el retículo sarcoplásmico se despolariza, el Ca2 + que contiene en sus cisternas terminales se descarga en el citoplasma y se une a la Troponina C (TpC), esta unión hace que se debilite el vínculo entre la troponina y la actina, permitiendo que la tropomiosina se desplace lateralmente y exponga el sitio activo donde la actina se une a la miosina. Por cada Ca2 + que se une a la troponina, se descubren 7 sitios de unión para la miosina. Es ahora cuando las cabezas de miosina se unen a los sitios de unión de la actina y una vez unidos, las cabezas de miosina actúan como bisagras moviéndose y arrastrando la cadena de actina.
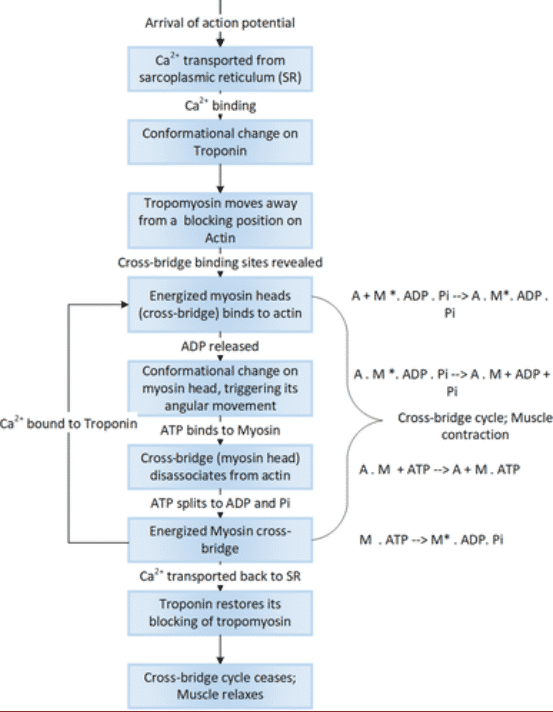
Figura 4 Proceso de contracción muscular11
La contracción muscular consiste esencialmente en la unión y separación de la cabeza de la miosina S-1 con los filamentos de la F-actina. La unión de la actina a la miosina genera cambios conformacionales dependientes del ADP o ATP, entre los que destaca la cabeza S-1. Estos cambios generan el llamado golpe de potencia, que dirige el movimiento de los filamentos de actina a través de los filamentos de miosina. La energía de este proceso es suministrada por el ATP, que es hidrolizado a ADP y Pi; sin embargo, el golpe de potencia se produce como resultado de los cambios de conformación cuando el ADP se desprende de las cabezas de miosina11.
Contracción muscular
3.1. Molecular
La miosina es una proteína hexamérica que contribuye en un 55% al peso de la proteína muscular y está compuesta por un extremo fibroso de dos hélices intercaladas, cada hélice tiene una cabeza globular unida a cada extremo. El hexámero consiste en un par de cadenas pesadas y dos cadenas ligeras diferentes (esenciales y reguladoras; ELC y RLC). Cada RLC tiene un sitio de unión específico para la incorporación de grupos de fosfatos. La fosforilación de la RLC es catalizada por la enzima miosina cinasa de cadena ligera, que se activa cuando se liberan moléculas de Ca2 + del retículo sarcoplásmico durante la contracción muscular12-15.
Una vez que el calcio llega al citoplasma de la célula muscular, se une a una proteína llamada calmodulina, formando un complejo de calcio y calmodulina16. El complejo de calcio y calmodulina activa una enzima llamada miosina cinasa de cadena ligera. Esta enzima cataliza la incorporación de grupos de fosfatos a las cadenas ligeras de la miosina, fosforilándolas y alterando el estado de los puentes cruzados según el estado de fosforilación13,15.
Hay abundantes pruebas científicas que demuestran que el fenómeno fisiológico de la potenciación es el resultado de la fosforilación de las CLR17-19. Numerosos estudios han demostrado la existencia de una correlación significativa entre la fosforilación de las CLR y la magnitud de la potenciación4.
Se cree que la fosforilación de las RLC mejora las contracciones posteriores al alterar la estructura de la cabeza de la miosina20. La fosforilación de las RLC hace que las cabezas de miosina se alejen de la espina dorsal del filamento grueso, disminuyendo la distancia entre los filamentos finos y gruesos, aumentando así la velocidad a la que puede acercarse a la actina. También se ha demostrado que la fosforilación RLC hace que la interacción actina-miosina sea más sensible al Ca2 +20,21 mioplásmico. Además, la fosforilación de las CLR daría como resultado una mayor cantidad de conformación activa de puente cruzado. Esto podría explicar en parte el aumento tanto de la producción de fuerza como de la tasa de desarrollo de fuerza registrada en el músculo esquelético de los mamíferos después del estímulo de acondicionamiento 22,23.
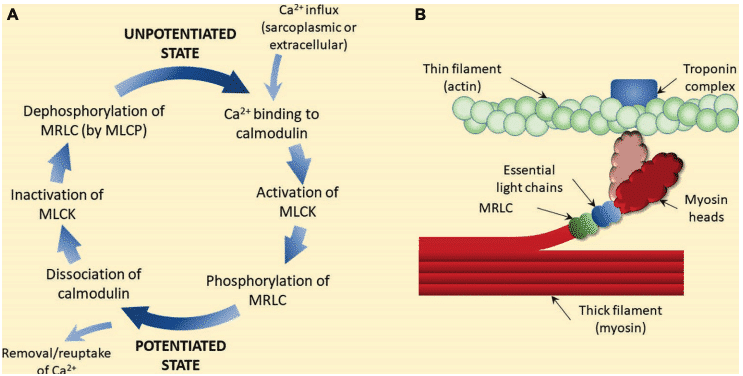
Figura 5 Mecanismo propuesto de potenciación post-activación (PAP)4
Por otra parte, las pruebas de que la fosforilación de las CLR es el principal mecanismo de la PAP no excluye la participación de otros mecanismos. Se ha demostrado que los cambios en la arquitectura muscular, relacionados específicamente con el ángulo de península (AP), son un potencial responsable de la generación de PAP. El AP está formado por los fascículos y la aponeurosis interna, que refleja la orientación de las fibras musculares en relación con el tejido conectivo. Por esta razón, cualquier cambio en ella afectará a la transmisión de la fuerza de los músculos a los tendones y huesos24,25.
3.2. Neuronal
Otros autores, sugirieron que la base fisiológica de la potenciación se basa en un efecto neural. Verkhoshansky propuso que cualquier estímulo previo a la acción motora, ya sea momentáneo o no, genera un «rastro en el sistema nervioso» que permite mantener los niveles de fuerza obtenidos durante cierto tiempo e incluso mejorarlos, lo cual fue el fundamento teórico del método de choque desarrollado por el mencionado autor. Esta mejora podría lograrse gracias a un mayor nivel de impulso neural voluntario.
Factores neuronales como el aumento del reclutamiento de unidades motoras de alto umbral, el aumento de la actividad de los músculos sinérgicos, el aumento de la actividad de los reflejos y la inhibición del aparato de Golgi podrían contribuir directa o indirectamente a la generación de PAP. El aumento de la actividad neuronal genera un mayor reclutamiento de unidades motoras de alto umbral, mejora la sincronización entre ellas, reduce la inhibición presináptica y genera un aumento de los impulsos del sistema nervioso central (SNC) hacia la periferia. El estímulo desencadenante (pre-contracción) aumenta la probabilidad de iniciar un potencial de acción cuya excitación generada puede durar varios minutos, aumentando los potenciales postsinápticos que conducen a una mayor generación de fuerza.
Sin embargo, por lo general no se observan aumentos en el EMG, incluso cuando se detectan claramente mejoras en el rendimiento funcional26-28 y, a pesar de los indicios de que los cambios neuronales pueden ser desencadenados por la actividad voluntaria, faltan pruebas que apoyen la teoría de que los aumentos del impulso neuronal al músculo contribuyen a los aumentos del rendimiento muscular voluntario complejo observados en muchos estudios.

PAP vs PAPE
Las contracciones musculares de alta intensidad, como las que se utilizan en el ejercicio máximo o en el levantamiento de pesas, requieren niveles máximos o casi máximos de activación muscular, por lo que la PAP no puede influir directamente en ellas. Sin embargo, se ha demostrado que la PAP puede modificar la relación entre la fuerza y la concentración de Ca+, por lo que puede aumentar la salida de fuerza en una liberación determinada de Ca+ (más fuerza con menos activación, que puede verse como un aumento del RFD) especialmente durante las contracciones breves en las que no se puede alcanzar la fuerza máxima22,29-31. Esto puede influir, pues, en el rendimiento durante las actividades intensivas de corta duración, como los saltos o las patadas32.
Las pruebas de la existencia de la PAP con respecto a las propiedades contráctiles son abundantes. Sin embargo, las pruebas de la PAP en las tareas relacionadas con el rendimiento han sido un tanto confusas debido a que los estudios no han logrado observar mejoras o, si se han observado, no pueden atribuirse a la PAP. Si bien algunos residuos de PAP podrían influir en la PAPE en sus primeras etapas después de una actividad de acondicionamiento, otros mecanismos deben desempeñar un papel importante en la PAPE.
La prueba de que la PAP ha sustentado la mejora del rendimiento (es decir, la PAPE) sólo puede aportarse cuando la variable de resultado medida durante una contracción voluntaria cambia con el mismo curso temporal que la respuesta clásica de la PAP, que se suele estimar mediante la medición de la fuerza máxima de contracción muscular4. Lamentablemente, este tipo de diseños de estudio no son abundantes.
Como ya hemos señalado, parece que la PAP es diferente de la PAPE y el término PAPE debería utilizarse preferentemente para identificar la posibilidad de que se produzca un fenómeno distinto del PAP clásico33. Sin embargo, un aspecto clave de PAPE que requiere una mejor comprensión es la falta de un efecto de mejora del rendimiento en los puntos temporales en los que se manifiesta PAP (3-4 min), ya que PAP debería ser la causa de PAPE.
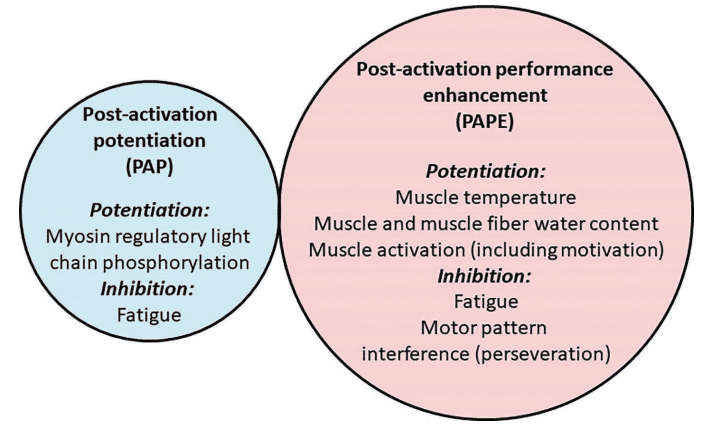
Figura 6 Posibles factores que influyen en la potenciación posterior a la activación (PAP) y en la mejora del rendimiento posterior a la activación (PAPE)4
Todavía no hay consenso científico. Sin embargo, la teoría dominante detrás de esta falta de efecto es la coexistencia de la potenciación y la fatiga25,34,35. Diferentes tipos de contracciones generarían efectos potenciadores en mayor o menor medida, y esto podría estar relacionado con el nivel de fatiga neuromuscular que producen. Después de una contracción muscular máxima o casi máxima, los músculos se encuentran en un estado de fatiga y potenciación simultáneamente. El estado potenciado permanece durante un período de tiempo determinado, tras el cual la fatiga disminuye y proporciona una «ventana» durante la cual el deportista puede obtener el beneficio ergogénico del estado potenciado, aunque parece que no es posible generalizar los períodos de recuperación 36,37. Además, la duración de la recuperación muestra una gran variabilidad entre individuos está asociada a numerosos factores como el nivel de fuerza, la experiencia de entrenamiento y la mio tipología36-38´, función muscular mediante mecanismos no relacionados con las CLR, como la acumulación de agua intracelular o los cambios en el flujo sanguíneo4.
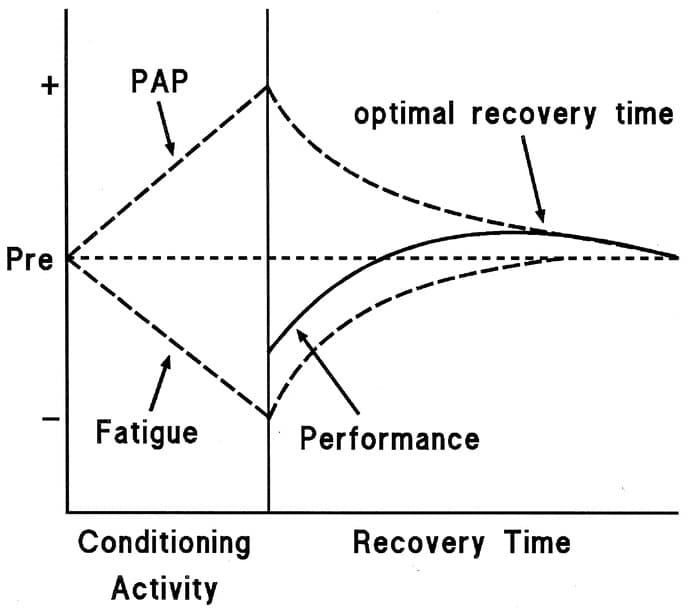
Figura 7 Coexistencia de PAP y Fatiga32
Otra posibilidad es que la falta de PAPE en los minutos posteriores a una actividad de acondicionamiento, a pesar de que la PAP es significativa, sea el resultado de un efecto de interferencia del patrón motor (a menudo llamado «perseverancia» ya que el patrón motor de una tarea persevera mientras se inicia una tarea similar y posterior). Sin embargo, también es probable que otros cambios fisiológicos aumenten la función muscular voluntaria en los puntos temporales en que se ha observado la PAPE, como los aumentos de la temperatura o la coordinación muscular (efectos de aprendizaje o motivación), o las mejoras de la función muscular mediante mecanismos no relacionados con las CLR, como la acumulación de agua intracelular o los cambios en el flujo sanguíneo4.
Los métodos que aprovechan el PAP, como el complejo o el contraste, probablemente sean útiles para mejorar la potencia mecánica y, por consiguiente, el rendimiento y/o el estímulo de entrenamiento de una actividad deportiva explosiva. Las pruebas sobre la aplicación práctica del PAP en actividades explosivas aún no son concluyentes, dada la diferencia antes mencionada entre el PAP y el PAPE. Parece que la respuesta de PAPE podría ser sólo el resultado de un buen, amplio y exitoso calentamiento.
Bibliografía
1.Lambert, M. I. General Adaptations to Exercise: Acute Versus Chronic and Strength Versus Endurance Training. in Exercise and Human Reproduction 93–100 (Springer New York, 2016). doi:10.1007/978-1-4939-3402-7_6.
2.Robbins, D. W. & Docherty, D. Effect of Loading on Enhancement of Power Performance Over Three Consecutive Trials. The Journal of Strength and Conditioning Research 19, 898 (2005).
3.MacIntosh, B. R., Robillard, M.-E. & Tomaras, E. K. Should postactivation potentiation be the goal of your warm-up? Applied Physiology, Nutrition, and Metabolism 37, 546–550 (2012).
4.Blazevich, A. J. & Babault, N. Post-activation Potentiation Versus Post-activation Performance Enhancement in Humans: Historical Perspective, Underlying Mechanisms, and Current Issues. Frontiers in Physiology 10, (2019).
5.Prieske, O., Behrens, M., Chaabene, H., Granacher, U. & Maffiuletti, N. A. Time to Differentiate Postactivation ‘Potentiation’ from ‘Performance Enhancement’ in the Strength and Conditioning Community. Sports Medicine 50, 1559–1565 (2020).
6.Lee, F. S. The cause of the treppe. Experimental Biology and Medicine 4, 22–23 (1906).
7.Bowditch, H. l] ber die Eigenth~ imlichkeiten der Reizbarkeit, welche die Muskelfasern des Herzens zeigen. Berichte Ober die Verhandlungen der k6nighch s~ chsischen Gesellschaft der Wissenschaften zu Leipzig. Mathemat.-Phys. Classe 23, 652–589.
8.Guttman, S. A., Horton, R. G. & Wilber, D. T. Enhancement of Muscle Contraction after Tetanus. Experimental Biology and Medicine 34, 219–221 (1936).
9.Brown, G. L. & von Euler, U. S. The after effects of a tetanus on mammalian muscle. The Journal of Physiology 93, 39–60 (1938).
10.Burke, R. E., Rudomin, P. & Zajac, F. E., III. The effect of activation history on tension production by individual muscle units. Brain Research 109, 515–529 (1976).
11.Mukund, K. & Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdisciplinary Reviews: Systems Biology and Medicine (2019) doi:10.1002/wsbm.1462.
12.Craig, R. & Woodhead, J. L. Structure and function of myosin filaments. Current Opinion in Structural Biology 16, 204–212 (2006).
13.Szczesna, D. et al. Phosphorylation of the regulatory light chains of myosin affects Ca2+sensitivity of skeletal muscle contraction. Journal of Applied Physiology 92, 1661–1670 (2002).
14.Manning, D. R. & Stull, J. T. Myosin light chain phosphorylation-dephosphorylation in mammalian skeletal muscle. American Journal of Physiology-Cell Physiology 242, C234–C241 (1982).
15.Davis, J. S., Satorius, C. L. & Epstein, N. D. Kinetic Effects of Myosin Regulatory Light Chain Phosphorylation on Skeletal Muscle Contraction. Biophysical Journal 83, 359–370 (2002).
16.Moore, R. L. & Stull, J. T. Myosin light chain phosphorylation in fast and slow skeletal muscles in situ. American Journal of Physiology-Cell Physiology 247, C462–C471 (1984).
17.Grange, R. W., Cory, C. R., Vandenboom, R. & Houston, M. E. Myosin phosphorylation augments force-displacement and force-velocity relationships of mouse fast muscle. American Journal of Physiology-Cell Physiology 269, C713–C724 (1995).
18.Houston, M. E., Lingley, M. D., Stuart, D. S. & Grange, R. W. Myosin light chain phosphorylation in intact human muscle. FEBS Letters 219, 469–471 (1987).
19.Sweeney, H. L., Bowman, B. F. & Stull, J. T. Myosin light chain phosphorylation in vertebrate striated muscle: regulation and function. American Journal of Physiology-Cell Physiology 264, C1085–C1095 (1993).
20.Szczesna-Cordary, D. Regulatory Light Chains of Striated Muscle Myosin. Structure, Function and Malfunction. Current Drug Target -Cardiovascular & Hematological Disorders 3, 187–197 (2003).
21.Baudry, S., Klass, M. & Duchateau, J. Postactivation potentiation of short tetanic contractions is differently influenced by stimulation frequency in young and elderly adults. European Journal of Applied Physiology 103, 449–459 (2008).
22.Vandenboom, R., Xeni, J., Bestic, N. M. & Houston, M. E. Increased force development rates of fatigued mouse skeletal muscle are graded to myosin light chain phosphate content. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 272, R1980–R1984 (1997).
23.Vandenboom, R., Grange, R. W. & Houston, M. E. Myosin phosphorylation enhances rate of force development in fast-twitch skeletal muscle. American Journal of Physiology-Cell Physiology 268, C596–C603 (1995).
24.Fukunaga, T., Ichinose, Y., Ito, M., Kawakami, Y. & Fukashiro, S. Determination of fascicle length and pennation in a contracting human muscle in vivo. Journal of Applied Physiology 82, 354–358 (1997).
25.Hodgson, M., Docherty, D. & Robbins, D. Post-Activation Potentiation. Sports Medicine 35, 585–595 (2005).
26.Hough, P. A., Ross, E. Z. & Howatson, G. Effects of Dynamic and Static Stretching on Vertical Jump Performance and Electromyographic Activity. Journal of Strength and Conditioning Research 23, 507–512 (2009).
27.Seitz, L. B., Trajano, G. S., Dal Maso, F., Haff, G. G. & Blazevich, A. J. Postactivation potentiation during voluntary contractions after continued knee extensor task-specific practice. Applied Physiology, Nutrition, and Metabolism 40, 230–237 (2015).
28.Mina, M. A., Blazevich, A. J., Giakas, G., Seitz, L. B. & Kay, A. D. Chain-loaded variable resistance warm-up improves free-weight maximal back squat performance. European Journal of Sport Science 16, 932–939 (2016).
29.Tillin, N. A. & Bishop, D. Factors Modulating Post-Activation Potentiation and its Effect on Performance of Subsequent Explosive Activities. Sports Medicine 39, 147–166 (2009).
30.Vandenboom, R., Gittings, W., Smith, I. C., Grange, R. W. & Stull, J. T. Myosin phosphorylation and force potentiation in skeletal muscle: evidence from animal models. Journal of Muscle Research and Cell Motility 34, 317–332 (2013).
31.Vandenboom, R., Grange, R. W. & Houston, M. E. Threshold for force potentiation associated with skeletal myosin phosphorylation. American Journal of Physiology-Cell Physiology 265, C1456–C1462 (1993).
32.Sale, D. G. Postactivation Potentiation: Role in Human Performance. Exercise and Sport Sciences Reviews 30, 138–143 (2002).
33.Cuenca-Fernández, F. et al. Nonlocalized postactivation performance enhancement (PAPE) effects in trained athletes: a pilot study. Applied Physiology, Nutrition, and Metabolism 42, 1122–1125 (2017).
34.Fowles, J. R. & Green, H. J. Coexistence of potentiation and low-frequency fatigue during voluntary exercise in human skeletal muscle. Canadian Journal of Physiology and Pharmacology 81, 1092–1100 (2003).
35.Rassier, D. E. & MacIntosh, B. R. Coexistence of potentiation and fatigue in skeletal muscle. Brazilian Journal of Medical and Biological Research 33, 499–508 (2000).
36.Gołaś, A., Maszczyk, A., Zajac, A., Mikołajec, K. & Stastny, P. Optimizing post activation potentiation for explosive activities in competitive sports. Journal of Human Kinetics 52, 95–106 (2016).
37.Naclerio, F. et al. Effects of Three Different Conditioning Activity Volumes on the Optimal Recovery Time for Potentiation in College Athletes. Journal of Strength and Conditioning Research 29, 2579–2585 (2015).
38.Chen, Z.-R., Lo, S.-L., Wang, M.-H., Yu, C.-F. & Peng, H.-T. Can Different Complex Training Improve the Individual Phenomenon of Post-Activation Potentiation? Journal of Human Kinetics 56, 167–175 (2017).